nucleotides and nucleic acids
- 中心法则
约90%的DNA都会转录成RNA,但是会翻译成蛋白质的很少。
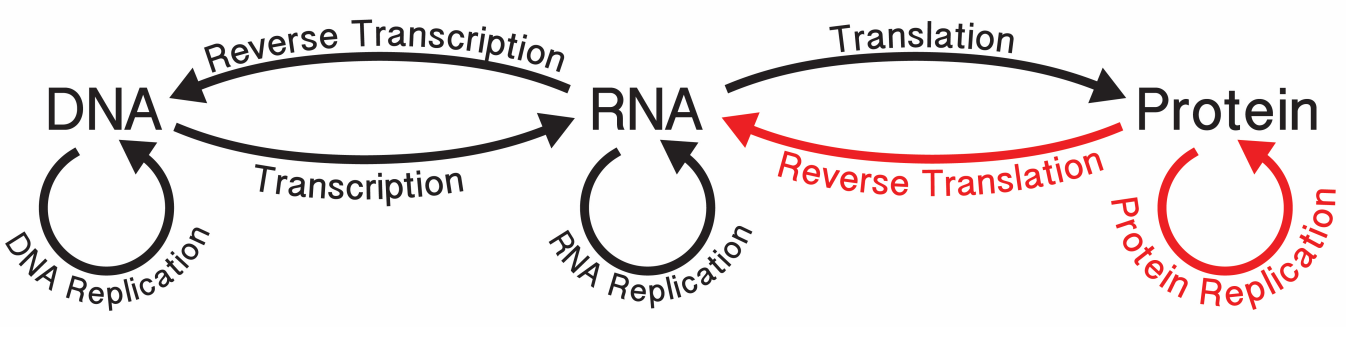
上图中红色部分为暂时无法通过生物体实现的部分 - 核酸研究的发展史
……原核生物的操纵子发现……
核酸的研究发展史
1865年Mendol 在布吕恩自然研究会上报告了他的豌豆杂交试验结果。
- 1868年Fridrich Miescher首次从脓细胞中提取出核素
- 1889年Altmann提出核酸的概念
- 1928年格里菲斯对小鼠进行了肺炎双球菌的注射感染实验
- 1944年Avery进一步分别提取出肺炎双球菌的DNA,RNA,蛋白质对细菌进行R菌转化实验,确定了DNA是转化因子(遗传物质)
- 1952年Hershey和Chase使用放射性同位素标记法用T2噬菌体对大肠杆菌进行了转染实验确认了DNA是遗传物质。
- 富兰克林用获得了DNA晶体衍射照片
- 1953年Watson和Crick在富兰克林研究的基础上发现了DNA的双螺旋结构。
- 1958年Crick提出“中心法则”
- 1961年Jacob和Monod提出操纵⼦学说
- 1966年Nirenberg等破译了遗传密码。
- 1970年 Temin和Baltimore发现逆转录酶,补充了中⼼法则。
- DNA重组技术被发明推广
- 1975/76年 Sanger和Maxam/Gilbert先后建立第一代DNA测序⽅法。
- 1985年 Mullis发明PCR技术。
- 1990年 美国启动⼈类基因组计划(HGP)。
- 1999年 中国⼈类基因组计划启动。
- 2003年 美英法德中日西等国共同完成⼈类基因组计划。
- 核酸的定义、功能、分类(DNA、RNA)、结构、分布(对于真核生物存在于线粒体,细胞核和叶绿体)
化学组成和一级结构
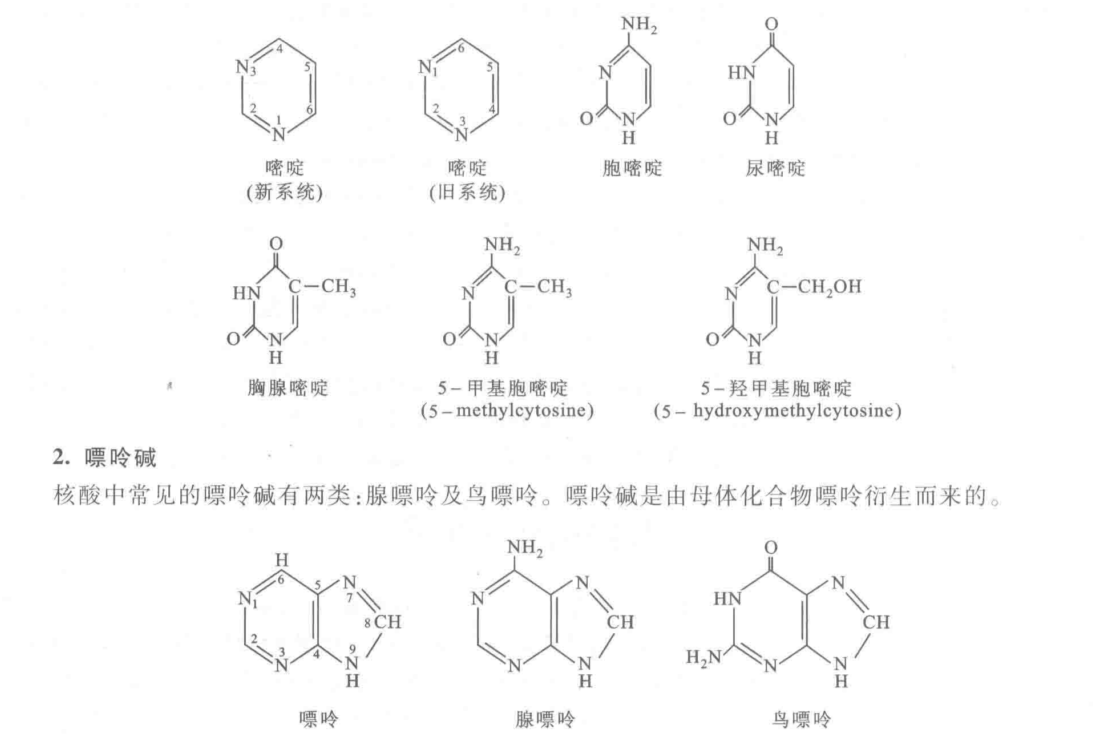
- 碱基的结构(要求记住碱基的结构)
- 核糖核苷酸简写AMP,CMP,TMP,GMP,脱氧前加d,例:dAMP[1]
- 核苷酸衍生物:
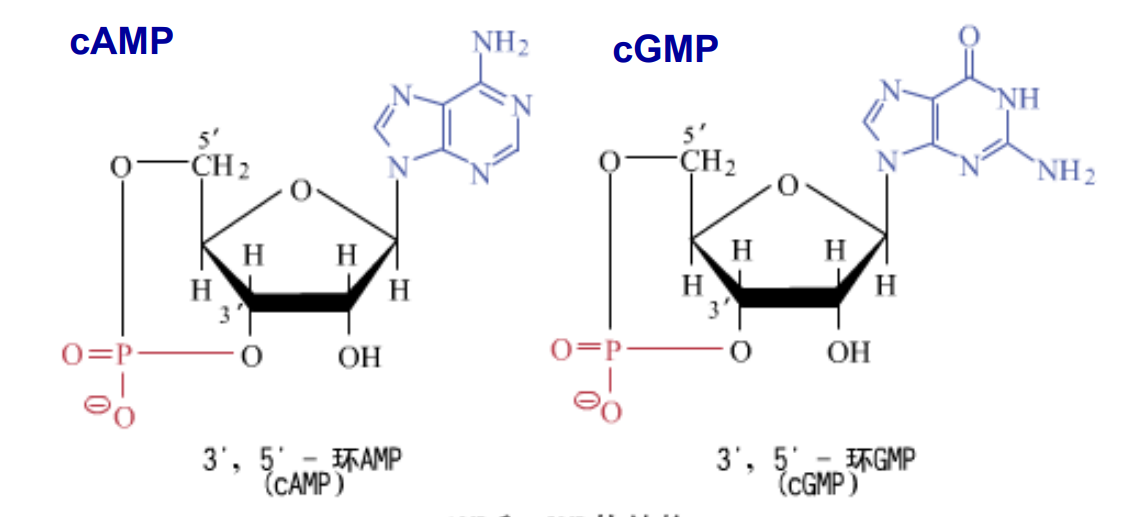
- 环化核苷酸 cNMP[2],磷酸基团与3‘位的羟基脱水形成共价键。
- 一些化学修饰如羟基、甲基等可能存在于核苷酸上···
- 磷酸基团的错位,通常情况下磷酸基团位于核糖的5’位置,但同时也存在2‘,3’位置的核苷酸。特别的,2‘,3’-环核苷酸是很多关键核酸反应的中间过渡态。
- 特殊的核苷酸:次黄嘌呤核苷酸,可与A、U、C形成非Watson-Crick配对。[3]
- 核糖的一号碳与碱基通过糖苷键(C-N)连接
- 多聚脱氧核苷酸(DNA链)具有方向性,方向是5‘(磷酸方向)到3’。
- 书写方法
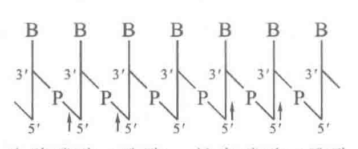
- 小核酸片段称为寡核苷酸。(<50bp)
- 与蛋白质相比,其元素组成不含S而含P,且P的含量较为稳定,约为9%-11%
- 一级结构:碱基的序列(核苷酸的组成与排列方式)
核苷酸的生物学应用
- 参与中心法则中各反应的各关键反应步骤;参与糖和磷脂的合成
- NTP(ATP)是生物体内能量硬通货
- 是构成多种辅酶的重要成分:NAD、NADP、FAD、FMN、CoA
- 参与细胞的代谢与调节(cAMP、cGMP)
DNA(RNA)的空间结构
两条脱氧核苷酸链通过氢键互补配对,随后亲水的脱氧核糖和磷酸骨架形成双螺旋结构将疏水的碱基包裹在内。
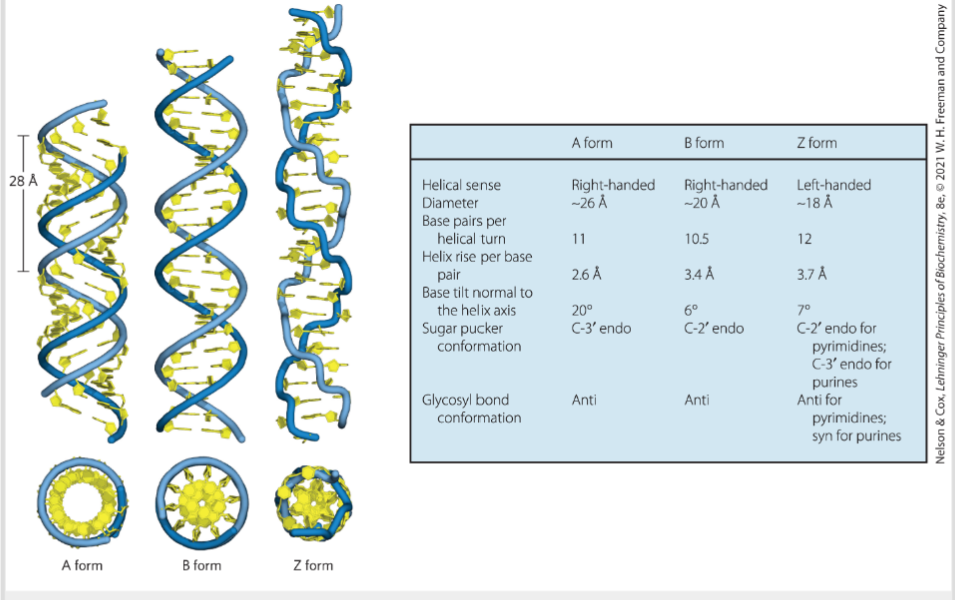
- DNA双螺旋结构: 分为A,B,C,Z型,还分为线性环状等若干种.
- B型DNA为自然状态下最常见的.
- A型DNA是在缺水的情况下提取出的;自然状态下RNA多为A型, 而DNA,RNA杂合形成的双链更像A型。A型较粗,螺距较小,约为2.8nm(相对于B型)
- Z-DNA为左手型骨架走向呈锯齿状(Zig-zag),C-G含量较多; 这种螺旋可能在基因表达或遗传重组中起作用。
- C-DNA,在44%-46%相对湿度的锂盐中的构型.(螺距3.1nm)
- 碱基对间存在大
键,碱基等特异性基团大多都分布在大groove一侧。 
- 氢键使两条链紧密结合,
键(碱基堆积力)和疏水作用力使DNA呈双螺旋状。 - RNA通常认为是单链,尽管可能有局部配对。
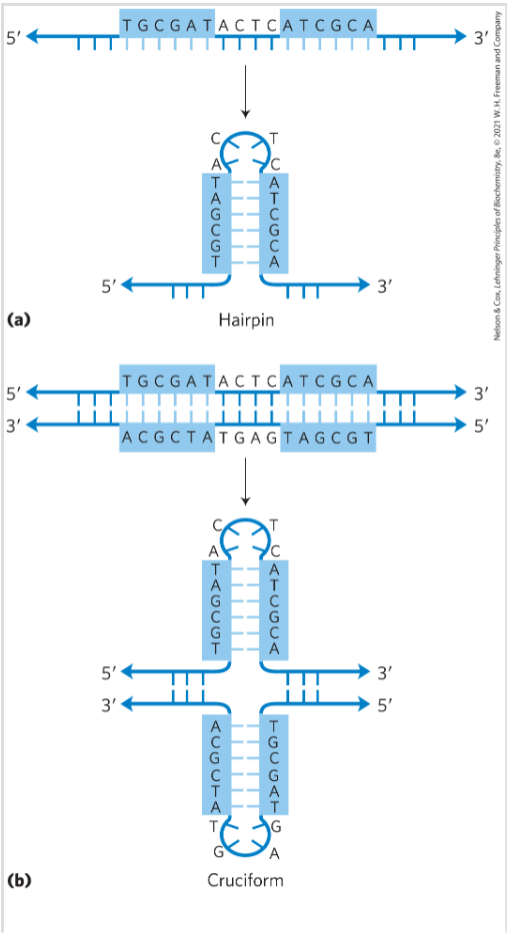
- DNA的发夹结构(单链自身局部互补);DNA的十字架结构(形成于DNA的回文结构,两个发夹结构,即双链的DNA两条单链各自出现了自身局域互补配对)
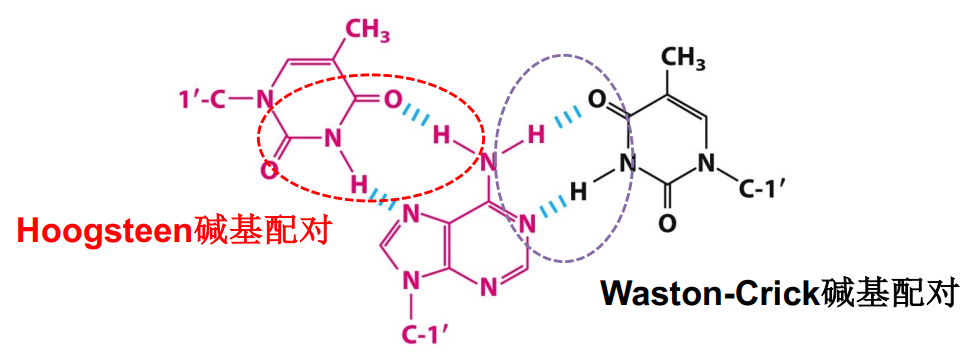
- 特殊的三链结构H-DNA:插入到major groove中,C,T含量高,且通常在酸性条件下形成。Hoogsteen氢键配对(与Watson-Crick区别)
- 四链结构,又称为G-四链体,通常连较短,每一层都是4个G,需要特殊序列。
- 真核生物DNA3'末端(端粒)是富含GT的重复序列,因而自身成为了四链结构.
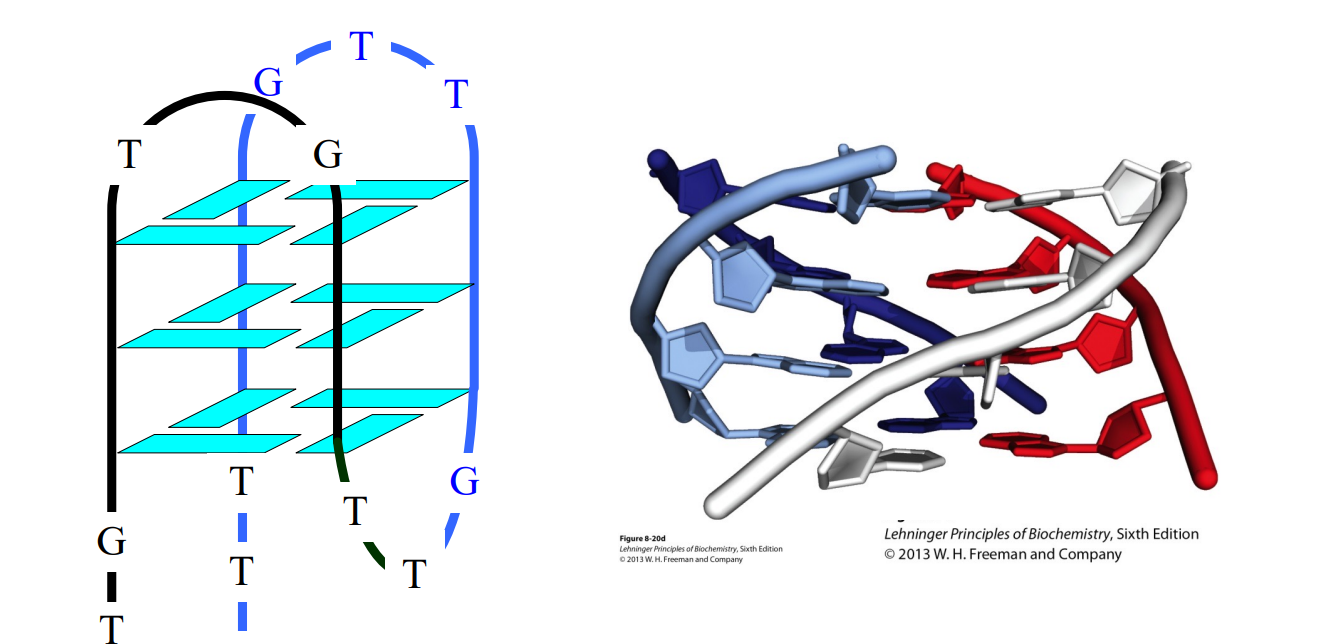
- 真核生物DNA3'末端(端粒)是富含GT的重复序列,因而自身成为了四链结构.
核苷酸与核酸的理化性质
- 天然粘性很高,变性后粘性急剧下降
- 高分子溶液的粘度往往要比球状分子的粘度要大.
#核酸的溶解性
- 高分子溶液的粘度往往要比球状分子的粘度要大.
- 主要受溶液盐浓度影响
- 盐浓度升高DNP的溶解度也升高
- DNP和RNP在盐溶液中的溶解度不同
- DNA为白色纤状固体,RNA为白色粉末.DNA溶于苯酚,RNA不溶.两者均不溶于乙醇乙醚等有机溶剂.
#核酸的沉降特性和密度特征
- 核酸的密度与其构象有关,超螺旋DNA的密度远大于一般线性的DNA分子.
- 利用不同DNA的浮力密度不同以分离不同DNA.(超高速离心)
#核酸的酸碱性
两性电解质,但偏酸性.
#核酸分子的紫外吸收的能力
- DNA与RNA在260nm均有强吸收,可以据此分辨DNA,RNA,单链DNA,寡核苷酸。
- DNA变性时有增色效应,复原后减色。
- A260:A280=1.8;双股DNA偏高则可能是RNA污染,偏低可能是蛋白和酚类污染.
#DNA的变性
本质是:对双链氢键和碱基堆积力的破坏,不改变一级结构(同蛋白质一样)
变形因素:热、酸、碱、尿素,胍(gua,一声)等有机物质
G-C含量越高,解链温度越高。
退火温度通常为解链温度(TM)-25℃
#复性过程
首先成核:形成局部双链的过程;紧接着拉链式快速变回双螺旋。
影响因素:温度、时间、DNA浓度
降温太快,温差太大都不利于复性。
#DNA序列的复杂性
是指一剪断的DNA制剂以核苷酸对数计的最长的非重复序列的长度。例如,…ATATAT…是dAT的重复共聚物,它的复杂性是2;…AGTCAGTC…是dAGTC的重复共聚物,它的复杂性是4.
对于那些不含重复序列的生物来说,其复杂性可以用来表示基因组的大小。
- 简单重复序列较多的DNA分子更容易在复性过程中进行互补配对。
#Southern/Northern/Western-Blot
DNA-RNA-Pro
核酸的研究方法
- 分离核酸的一般原则
- 保证DNA一级结构的完整性:避免物理的剪切包括震荡,搅拌等
- 排除其他分子污染
- 一般步骤:破碎细胞-除去蛋白质,多糖,脂类生物大分子-沉淀核酸-去除盐类,有机溶剂-纯化干燥-溶解
- 避免核酶在分离纯化过程中与样品接触,尤其是RNA!(DEPC,RNase的不可逆抑制剂)
- DNP 易溶于浓盐溶液,不溶于生理盐水。
- 蛋白质会被酚类物质破坏结构
- ···
- 浓度测定办法:定糖法-紫外吸收法-定磷法
- Nanodrop或者Thermo紫外吸收法直接测定核酸的浓度
- 定糖法:
- DNA分子中的脱氧核糖在冰醋酸或浓硫酸存在 下可与二苯胺反应生成兰色化合物,该化合物可在595-620nm 波长下进行比色测定。
- 分子中的脱氧核糖在冰醋酸或浓硫酸存在 下可与二苯胺反应生成兰色化合物,该化合物可在595-620nm 波长下进行比色测定。
- 定磷法:RNA和DNA中都含有磷酸,可以进行磷的测定。纯的核酸中含磷量在9.5%左右,因此,测出核酸中的磷含量就可计算核酸的含量。
- 测定时先将核酸用强酸消化成无机磷酸, 后者与定磷试剂中的钼酸反应生成磷钼酸,在经过还原作 用而生成蓝色的复合物,最后在650-660nm进行比色测定。
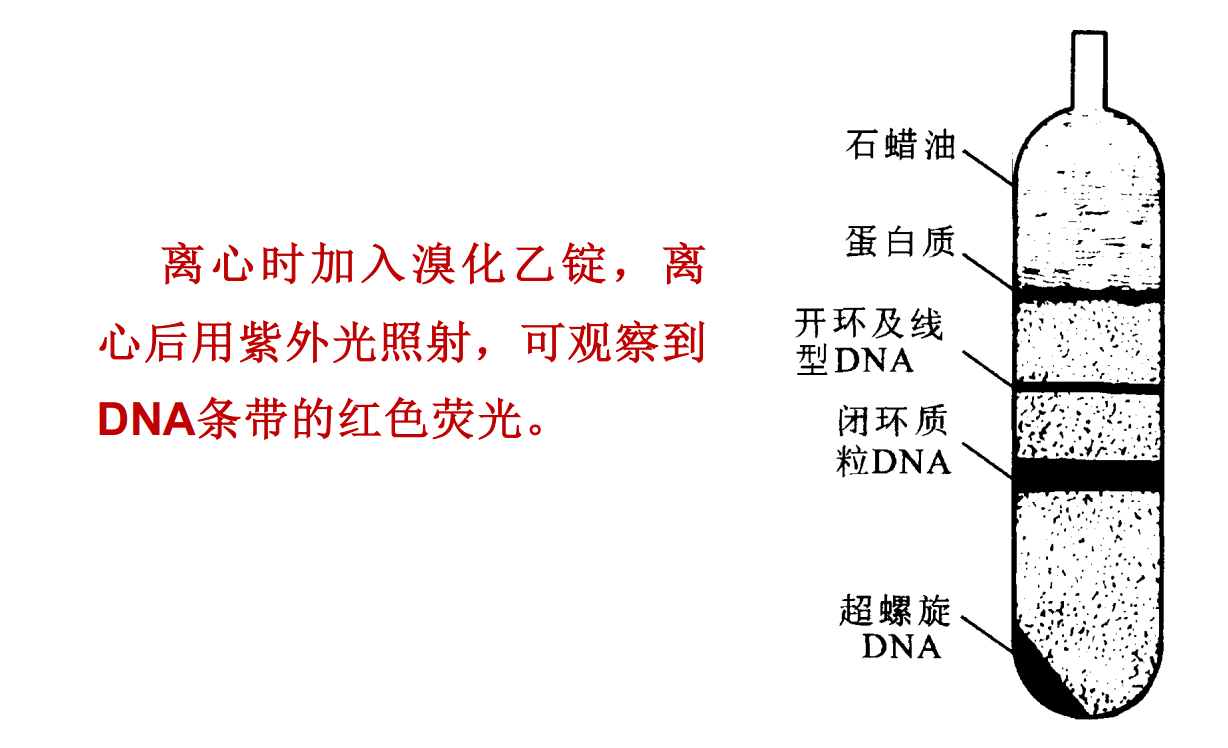
- 超速离心
- 作用:
- 测定核酸的浮力密度;(DNA溶于8.0mol/L的CsCl)
- 测定DNA分子中的G、C含量;(G-C多的DNA密度大)
- 测定溶液中核酸的构象,
- 核酸经染料—氯化铯密度梯度离心以后,在离心管里,沉降的速度由大到小依次为:超螺旋DNA、闭环质粒DNA、开环及线型DNA,若样品中含有蛋白质,蛋白质沉降速度最小。
- 核酸的制备
- 凝胶电泳
- 琼脂糖
- 核酸分子的大小。迁移率与分子量的对数成反比。
- 胶浓度。胶浓度大则网孔小,核酸的迁移率变小。
- 超螺旋的DNA迁移率 > 线性DNA > 开环DNA
- 聚丙烯酰胺(PAGE)
- 比琼脂糖凝胶电泳分辨率高,可分辨长度差1个核苷酸的两个DNA分子。
- 只能分离小分子量的DNA。
- 琼脂糖
- 染色
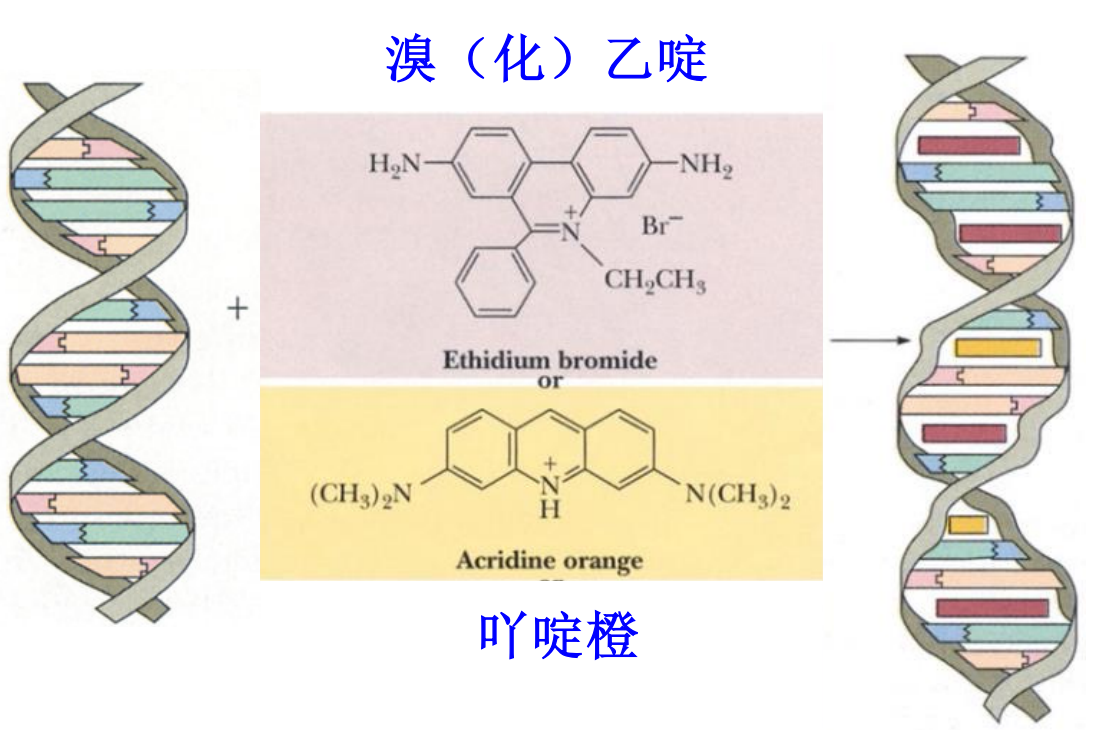
- 溴乙锭染色(致癌)
- 银染(贵)(PAGE)